Interleukin-1 in Febrile Infection-Related Epilepsy Syndrome
Article information

Abstract
Febrile infection-related epilepsy syndrome (FIRES) characteristically affects previously healthy children, who experience a sudden and explosive onset of super-refractory status epilepticus preceded by febrile infection and accompanied by fulminant neurogenic inflammation. FIRES, however, can affect individuals of all ages and is a subcategory of new-onset refractory status epilepticus. This definition of FIRES excludes febrile status epilepticus in infants. FIRES is a rare type of epileptic encephalopathy with rapidly progressive onset of seizures and a devastating prognosis, as drug-resistant epilepsy often follows without a latency period. Although the exact pathogenesis of FIRES has not been elucidated, a functional deficiency in the endogenous interleukin-1 receptor antagonist has been implicated in a genetic predisposition to FIRES. Dysregulation of the interleukin-1β–interleukin-1 receptor 1 (IL-1β–IL-1R1) signaling pathway appears to be involved in the pathogenesis of FIRES. In this review, the authors summarize the definition of FIRES, IL-1β–IL-1R1 signaling, the nucleotide-binding oligomerization domain the NLRP3 inflammasome, and IL-1 targeted therapy for FIRES.
Introduction
Febrile infection-related epilepsy syndrome (FIRES), a subcategory of new-onset refractory status epilepticus (NORSE), is a rare and devastating neurological condition with a high incidence of mortality and poor neurocognitive outcomes [1]. The estimated incidence of FIRES is one in a million, and its prevalence is one in 100,000 [2]. The historical mortality rate for FIRES during the acute phase is 9% to 18%. Additionally, only 18% of children maintain normal cognitive function in the acute phase, while more than 90% develop refractory epilepsy that requires lifelong treatment [1-3].
FIRES tends to affect children and young adults suddenly and explosively following a recent febrile illness. It often begins as new-onset intermittent seizures. The seizures subsequently increase in frequency and duration, and then progress to refractory status epilepticus (RSE) over 2 to 7 days with minimal response to anti-seizure medications (ASMs). Common features reported on electroencephalography in children with FIRES include (1) extreme delta brush; (2) a gradual increase in seizure burden; and (3) focal seizure activity, with an onset low-amplitude fast (>10 Hz) activity that evolves and shifts from one hemisphere of the brain to the other, eventually ending in the contralateral hemisphere [4]. Eventually, seizure activity weakens, and consciousness of varying degrees is gradually restored in surviving patients. Unfortunately, surviving patients often develop severe cognitive decline and chronic refractory epilepsy [5,6].
A crucial role of interleukin-1β (IL-1β) in FIRES has been demonstrated in case reports describing the effective clinical use of anakinra, a recombinant IL-1 receptor antagonist (IL-1Ra) [1,7]. Moreover, the levels of endogenous IL-1Ra and IL-1β were elevated both in the serum and cerebrospinal fluid (CSF) of patients with FIRES (n=7) relative to healthy controls (n=14, n=7 for CSF) [7]. Specifically, CSF IL-1Ra levels were markedly elevated in patients with FIRES [7]. These results suggest that the IL-1β–IL-1 receptor type 1 (IL-1R1) signaling pathway may play an important role in the pathogenesis of FIRES, and a better understanding of IL-1 in FIRES is needed to improve the treatment of FIRES patients.
Diagnosis of FIRES
A consensus-defining group convened and set standard definitions for NORSE and FIRES to unify clinicians and streamline research on NORSE and FIRES in 2018 [8]. NORSE was defined as a clinical presentation, not a specific diagnosis, involving the new-onset of a persistent refractory state of epilepsy without clear acute or active structural, toxic, or metabolic causes in patients without active epilepsy or other pre-existing associated neurological disorders. The International League Against Epilepsy defines FIRES as a subcategory of NORSE that requires a prior febrile infection, with fever starting between 2 weeks and 24 hours prior to the onset of RSE, with or without fever at the onset of status epilepticus [8]. FIRES includes all ages and excludes most cases of febrile status epilepticus in young children (prolonged febrile convulsions). This is because febrile seizures usually occur in children whose fever begins 24 hours before the seizure or is recognized only after the onset of the seizure [9]. Although this definition may include some unusual cases of “febrile status epilepticus” in which a fever persists for more than 24 hours and status epilepticus is refractory, this situation may reflect pathophysiology similar to that of other cases presenting with NORSE, despite being on the mild end of the spectrum. Unlike NORSE, FIRES affects slightly more boys than girls, and the median age of onset is approximately 8 years [8,10].
Basic Pathogenesis of FIRES: Blood-Brain Barrier Disruption and Neuroinflammation
The exact pathogenesis of FIRES has yet to be elucidated, although fulminant inflammation in the brain has been implicated [2,11]. Neurogenic inflammation has been proposed to occur as an inflammatory response in cells within the central nervous system (CNS), not only in neurons and glia, but in perivascular cells of the blood-brain barrier (BBB) [12]. A nonspecific, febrile infectious process occurs within 2 weeks before seizure onset. However, no single causative pathogen has been identified [5]. Rather, it is likely that a nonspecific infection, not causal to the syndrome, triggers an inflammatory cascade [5]. The hypothesis, supported by increased CSF levels of inflammatory molecules and the therapeutic response to immunomodulatory therapy, is that progressive febrile infection induces an inflammatory response that reduces the seizure threshold, stimulating the brain of predisposed individuals [1]. Days to weeks after the febrile infection, the reduced threshold favors the precipitation of seizures, triggering a massive neurogenic inflammatory response [1]. These responses contribute to seizure recurrence and status epilepticus [1]. Low rates of multiple autoantibodies, including glutamine decarboxylase, glutamate receptor subtype 2, glutamate receptor subtype 3, anti-voltage-gated potassium channel complex, and neuropil, have been detected in FIRES cases [1,5,13]. However, these rare positive findings appear to reflect nonspecific, secondary epiphenomena associated with BBB breakage [5,13].
IL-1 and Its Relevance: IL-1, IL-1R1, IL-1Ra, and NLRP3
IL-1 is the master cytokine of local and systemic inflammation. IL-1Ra is an endogenous competitive antagonist of IL-1R1. This receptor transduces cellular signals upon agonist activation. Over 100-fold molar excess of IL-1Ra is required to inhibit IL-1 activity efficiently [14]. The production of IL-1 is stimulated by exogenous Toll-like receptor (TLR) agonists or endogenous cytokines such as tumor necrosis factor-α [15]. Both IL-1α and IL-1β induce themselves, and this self-sustaining induction of IL-1 leads to autoinflammation. To prevent unwanted release and inflammatory runaway, IL-1β is synthesized as an inactive precursor that is activated following proteolytic cleavage by the intracellular cysteine protease, caspase-1 [16]. In turn, the activation of caspase-1 requires the oligomerization and assembly of the “inflammasome,” a complex of intracellular proteins [16]. Once activated, caspase-1 cleaves the N-terminal amino acid of the inactive IL-1β precursor, enabling the release of this cytokine's biologically active form. The assembly and activation of the inflammasome represent an important safety mechanism to prevent the deregulated release of IL-1β. Unlimited activation of caspase-1 and secretion of IL-1β induce systemic and multi-organ sterile inflammation, a hallmark of autoinflammatory diseases [17].
Inflammasomes are cytoplasmic high-molecular-weight protein platforms of caspase-1 activation in response to microbial invasion and danger signals [18]. Inflammasomes consist of the nucleotide-binding oligomerization domain-like receptor (NLR) family, the adapter apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC), and the effector protease caspase-1. The formation of these protein complexes results in the activation of caspase-1, which is involved in the maturation of the proinflammatory cytokines IL-1β and IL-18 into biologically active forms and the cleavage of gasdermin D (GSDMD) to promote pyroptotic cell death (pyroptosis) [18,19]. Among inflammasomes, the nucleotide-binding oligomerization domain (NOD)-, leucine-rich repeat (LRR)-, and pyrin domain (PYD)-containing protein 3 (NLRP3) inflammasome has been studied extensively and has been found to be activated by a broad spectrum of stimuli (Fig. 1). In general, NLRP3 inflammasome activation is regulated through a two-step process, with priming at the transcriptional and posttranslational levels (signal 1) and assembly by multiple pathways in response to a variety of exogenous pathogen-derived or endogenous danger molecules (signal 2) [18,19].
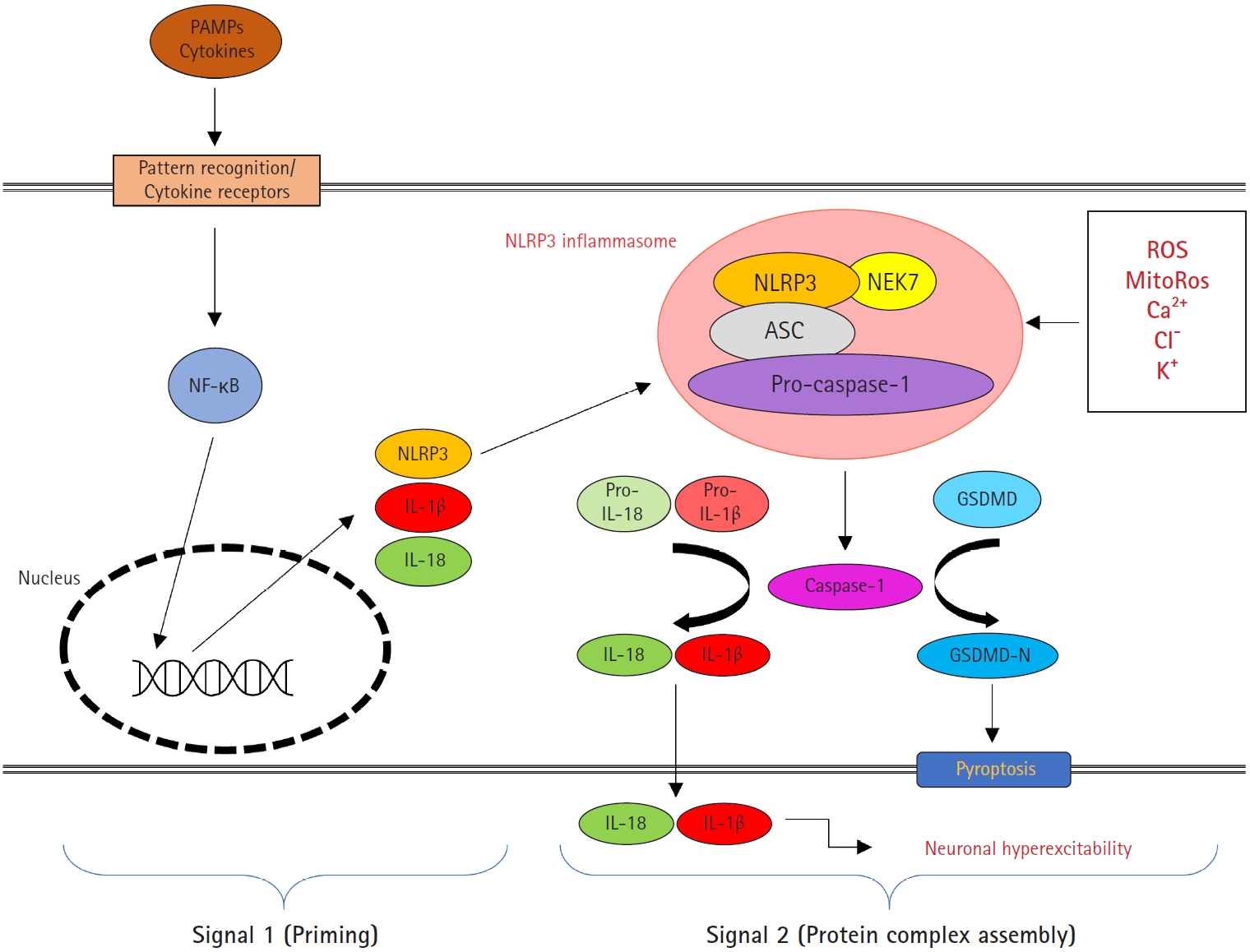
Schematic overview of the nucleotide-binding oligomerization domain (NOD)-, leucine-rich repeat (LRR)-, and pyrin domain-containing protein 3 (NLRP3) inflammasome. NLRP3 inflammasome activation involves two steps: signal 1 (priming) and signal 2 (protein complex assembly). Signal 1 is triggered by pattern recognition receptor signaling or cytokines, leading to the transcriptional activation of NLRP3 inflammasome components via the nuclear factor kappa B (NF-κB) pathway. Licensing the NLRP3 protein is essential for the priming step of the NLRP3 inflammasome. The NLRP3 inflammasome consists of pro-caspase-1, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC), NLRP3, and NIMA-related kinase 7 (NEK7). Various pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs) induce the activation signal 2 by reactive oxygen species (ROS), mitochondrial reactive oxygen species (mitoROS), Ca2+, Cl-, and K+. The NLRP3 complex leads to autoproteolytic activation of caspase-1 and triggers the cleavage of the interleukin 1β (IL-1β) and IL-18 and the pyroptotic substrate gasdermin D (GSDMD). IL-1β subsequently increases neuronal excitability.
IL-1 and Seizures
The activation of the IL-1β–IL-1R1 signaling pathway in human refractory epilepsies in both children and adults has been documented by immunohistochemical and biochemical studies performed in surgically resected epileptogenic lesions [20-24]. Similar findings have been reported in both immature and adult animal models of acute symptomatic seizures, febrile and non-febrile status epilepticus, and models of acquired epilepsies, absence epilepsy, and progressive myoclonic epilepsy [25-28]. In particular, the activation of this signaling pathway in animals significantly contributes to seizure generation in both acute and chronic models and is involved in epileptogenesis provoked by status epilepticus [29-31]. The activation of the IL-1 signaling pathway, a central part of the neuroinflammatory response, in chronic epileptogenic tissue suggests that this signaling is inefficiently controlled [32,33]. Indeed, IL-1Ra, which is the critical molecule modulating the effects of IL-1β, is expressed to a lower extent than IL-1β in epileptic foci from drug-resistant epilepsy patients and in animal models [21,34]. There is also a lower level of IL-1Ra in the blood of children with febrile seizures than in children with fever who do not have seizures [35]. Therefore, it is likely that the IL-1Ra:IL-1β ratio determines the seizure threshold. NLRP3 is also crucial in the IL-1β–IL-1R1 signaling pathway, and increased expression of NLRP3 has been reported in febrile seizure and temporal lobe epilepsy [36-38].
IL-1 and FIRES
FIRES may be considered a postinfectious immune system dysregulation in healthy but vulnerable individuals. Although no consistent single-gene mutations have been reported in patients with FIRES, a genetic predisposition for inflammation may exist. A potential link to the cytokine pathway was identified in patients with FIRES in 2016. A candidate gene analysis of FIRES in 19 Japanese children, focusing on polymorphisms of cytokine-associated genes, found a significant association between the tandem repeat frequencies of the RN2 allele of IL1RN and FIRES [39]. Mutations in this allele result in higher levels of IL-1β and lower levels of IL-1Ra. A potential imbalance of endogenous IL1-Ra and active IL-1β, reflecting an intrinsic functional deficiency, could lead to an opposing pathologic inflammatory state. Sakuma et al. [11] found that proinflammatory cytokines and chemokines were significantly elevated in FIRES patients, whereas T cell-associated and homeostatic cytokines were not [7]. Recent evidence has also suggested that circulating IL-1Ra in children with FIRES is less effective at blocking IL-1β signals than native IL-1Ra, as assessed in a cell-based reporter system, indicating a functional inefficiency in IL-1Ra inhibitory activity [7].
IL-1 Targeted Therapy
1. IL-1Ra blocker: anakinra
IL-1Ra is an endogenous member of the IL-1 family that binds to IL-1R1 and blocks the activity of IL-1α and IL-1β [40]. Anakinra is the recombinant human form of IL-1Ra and was first introduced in 1993. Anakinra currently has an established safety profile and known pharmacokinetics with a short half-life, and it is known to be effective in the CNS [41]. Anakinra is used to treat a variety of diseases, from common conditions such as rheumatoid arthritis, gout, and idiopathic pericarditis, to rare hereditary diseases such as hemophagocytic lymphohistocytosis [41,42]. Certain mutations in diseases such as familial Mediterranean fever and cryopyrin-associated periodic syndrome result in the uncontrolled release of active IL-1β, which clinically results in periodic fevers with systemic and local inflammation [14]. In phase II clinical studies evaluating the use of anakinra for neurological disorders, including acute stroke and traumatic brain injury, patients demonstrated tolerability of the drug even at exceptionally high doses [43,44].
2. The preclinical outcomes of anakinra, an IL-1R blocker, in seizure models
In a temporal lobe epilepsy model of systemic kainic acid (KA) injection in adult rats, intracerebral injections of IL-1Ra decreased seizure onset, the number of seizures, and the time in seizures by 50%. An altered balance between IL-1β and IL-1Ra appears to determine seizure threshold and contribute to ictogenesis [45]. Another study showed that adult mice overexpressing the IL-1Ra in astrocytes by 15-fold were intrinsically more resistant to seizures [46]. Further studies of anakinra, an Il-1R antagonist, in adult rodent models have demonstrated its effect on the incidence, severity, and duration of seizures [47,48]. Anakinra, which also mediates neuroprotection, inhibited seizures induced by bicuculline in an isolated guinea pig brain [49]. The anticonvulsive effect of anakinra was associated with a reduction in IL-1β expression in astrocytes and rescue of BBB permeability dysfunction [50]. Furthermore, the inhibition of IL-1β biosynthesis with the caspase-1 inhibitor VX-765 effectively reduced drug-resistant recurrent seizures in adult epileptic mice [51].
Since FIRES primarily affects children and young adults, it is relevant to highlight experimental studies involving immature animals. Anakinra reduced kindling epileptogenesis in immature rats promoted by lipopolysaccharide (LPS) [52]. Anakinra combined with a cyclooxygenase-2 antagonist given to young postnatal day (P) P21 rats after status epilepticus reduced the severity of subsequent epilepsy; the rats developed a mild form of epilepsy, with a reduction of spontaneous seizure frequency (up to 70%–90%) [53]. The study also observed reduced cell death and behavioral deficits. Immunomodulation afforded neuroprotection and rescued neurological comorbidities [53]. Activation of the IL-1–IL-1R1 axis has been demonstrated in forebrain astrocytes and neurons in immature P14–15 mice models of hyperthermia-induced status epilepticus [54]. The IL-1–IL-1R1 axis has been shown to contribute to both acute seizures and epilepsy development in these models of prolonged experimental febrile seizures [29,54]. Another model is the inflammatory challenge induced by the bacterial product LPS in immature rats [55]. Seizures were triggered later in life by a subconvulsive dose of chemoconvulsant, suggesting that the induction of seizures and long-term neurological sequelae involved inflammatory processes [55]. In particular, immature rats exposed to KA-induced seizures developed age-dependent neuroinflammation in the forebrain at about 2 weeks of age and approached the adult pattern by P21 [56]. This age-dependent pattern may be attributed to a dissociation between KA-induced seizures and activation of transcriptional factors that promote inflammation, such as activator protein-1 (AP-1) or nuclear factor kappa B (NF-κB). The rodent brain is able to exhibit seizure-induced neuroinflammation at an age comparable to school-aged children who are susceptible to developing FIRES [57].
3. Clinical use of anakinra, an IL-1R blocker
1) Case reports
In 2016, the first case of the use of anakinra to successfully treat a 32-month-old girl with FIRES was reported [58]. Her status epilepticus was previously unresponsive to anesthetic agents, midazolam, and phenobarbital. With prolonged treatment with anakinra (5 mg/kg twice daily, maximum; 200 mg) initiated 6 days after status epilepticus onset and multidisciplinary intervention, the patient showed no developmental or cognitive impairment, except for rare focal seizures. After the first case of anakinra use for FIRES, nine cases, in patients ranging from 32 months to 21 years of age, were presented in case reports [58-65]. Anakinra was mostly administered after various treatments, including anesthetics, ASMs, and anti-immunotherapy. The mean medication count before using anakinra was 8.9±2.6 (median, 10; range, 5 to 12). All patients were treated with a ketogenic diet (KD) and steroid therapy. Eight patients (8 /9, 89%) received intravenous immunoglobulin, and five patients (5/9, 56%) underwent plasmapheresis. Anakinra was administered at 5 to 20 mg/kg/day for 6 to 540 days after the onset of FIRES. Excluding a case with anakinra use in a chronic state (at 540 days) of FIRES [61], the mean starting time of anakinra was 26.3±13.2 days (median, 30; range, 6 to 43) after seizure onset [58-60,62-65]. Seven patients (7/9, 78%) reached >50% seizure reduction. There was one case of drug rash with eosinophilia and systemic symptoms (DRESS) (case no.1) [58]. Otherwise, there were no specific side effects due to anakinra. Four patients (4/9, 44%) recovered their cognitive function to near baseline before the onset of FIRES [58-65].
2) Cohort study
An international cohort study of 25 patients with FIRES treated with anakinra was published in 2020 [66]. The median age of the patients was 8 years (interquartile range, 5.2 to 11), and all were treated with anesthesia for seizure control prior to the start of anakinra. All children received at least four additional ASMs, with seven or more agents failing in 18 (72%) patients prior to anakinra initiation. Anakinra was started at a median of 20 days (interquartile range, 14 to 25) after seizure onset, with an initial median dose of 3.8 mg/kg/day (interquartile range, 3 to 5) and a final median dose of 5 mg/kg/day (interquartile range, 4 to 9). A seizure reduction of >50% at 1 week of anakinra treatment was observed in 11 of 15 patients (73%) with available seizure frequency data. The neurological outcomes were overall poor, and the differences associated with the degree of seizure control were not statistically significant due to the small sample size. A trend toward a more extended period of mechanical ventilation, intensive care unit (ICU) stay, and hospital length of stay was observed in patients without favorable seizure responses. In the evaluation of the Pediatric Cerebral Performance Category (PCPC), six patients had no or mild impairment (PCPC 1–2), six had moderate impairment (PCPC 3), and five had severe impairment or were in a vegetative state (PCPC 4–5). All surviving children had refractory epilepsy. Twelve children returned to school, and 10 required academic accommodations or special education classes. In that study, early anakinra initiation was associated with a shorter period of mechanical ventilation, shorter ICU and hospital lengths of stay, and possibly, seizure reduction. However, the authors could not ascertain the optimal therapeutic window for anakinra treatment [66].
3) Refractoriness to anakinra in FIRES
In the case reports, two out of nine patients were refractory to anakinra. Case no. 6 responded effectively to tocilizumab, an IL-6 receptor antagonist. The patients had elevated IL-6 levels in the CSF, and anakinra was switched to tocilizumab. The seizures improved, but behavioral dysregulation and inattention persisted [62]. Case no. 4 was resistant to various treatments, including deep brain stimulation of the centro-median thalamic nuclei, and remained in a vegetative state with frequent focal seizures (Table 1).

Summary of FIRES patients with anakinra usage
Fifteen patients were available in a cohort study with >50% seizure reduction data [66]. Four patients did not reach a 50% reduction in seizures after 1 week of anakinra treatment [66]. In various autoimmune diseases, the response to anakinra is usually evaluated within 1 to 3 months after initiation of treatment [67]. In FIRES, an evaluation at 1 week could be too short to determine the efficacy. The anakinra-refractory patients tended to show a longer duration of mechanical ventilation and ICU length of stay, although this difference was not statistically significant [66]. The prognosis at discharge was moderate disability, severe disability, vegetative state, or death [66]. It is difficult to determine the risk factors for poor response to anakinra due to small patient numbers. However, from the case series and cohort data, poor anakinra response was correlated with a poor prognosis.
4) Anakinra in the international consensus recommendations
The International NORSE Consensus Group published international consensus recommendations for the management of NORSE, including FIRES, in 2022 [68]. The consensus was made by a panel of 48 experts. They recommended treatment similar to the acute treatment of RSE during the initial 48 hours. First-line immunotherapy (corticosteroids, intravenous immunoglobulin G, or therapeutic plasma exchange) should be initiated within the first 72 hours of onset of status epilepticus. In cryptogenic NORSE/FIRES without clinical features of a specific autoimmune encephalitis syndrome, IL-1Ras or IL-6 blockers should be strongly considered as second-line immunological treatment [68].
4. Long-term use of anakinra
According to the case series (Table 1), four of nine patients continued anakinra until the time of case report publication [58,60,61,63]. Case no. 1 showed DRESS syndrome on day 22. After discontinuation of all drugs, the DRESS was improved. From day 54, anakinra was used again, and DRESS did not recur [58]. Two patients used anakinra for 22 and 210 days, respectively, and stopped it because of seizure reduction [59,65]. Due to ineffectiveness, the other two patients discontinued anakinra at 14 and 90 days of usage (Table 1) [60,62].
In 2022, the Paediatric Rheumatology International Trials Organization reported long-term safety data on anakinra in 306 patients with systemic juvenile idiopathic arthritis [69]. Among these 306 patients, 46%, 34%, and 28% had been treated for at least 12, 18, and 24 months, respectively. In total, 201 adverse events (AEs), mostly represented by infections, were reported for 509 patient-years (PY) with an overall incidence rate (IR) of 40 per 100 PY. Among 56 serious AEs (IR=11/100 PY), 23% were infections, and 20% were macrophage activation syndrome episodes. The IR of AEs was higher during the first 6 months of anakinra treatment, followed by decreasing IRs during long-term treatment. Treatment was discontinued in 76% of patients, most frequently in the first 6 months, because of lack of efficacy (43%), remission (31%), or AEs/intolerance (15%). No deaths or malignancies occurred during anakinra treatment [69]. Although similar safety data are not available for FIRES, anakinra might be considered safe in FIRES patients. Long-term follow-up research on anakinra use in FIRES is needed and may be possible via an international registry.
5. Ketogenic diet
KD is an established treatment for drug-resistant epilepsy and is a preferable option for FIRES over a prolonged drug-induced coma [5,70]. Ketone bodies and reactive oxygen species (ROS) modulation could decrease IL-1β levels by inhibiting NLRP3 expression. Beta-hydroxybutyrate (BHB) is the main ketone body resulting from the oxidation of fatty acids in the liver. Several studies have concurred in favor of a direct anti-inflammatory role for BHB by direct inhibition of NLRP3 inflammasome assembly [71,72]. BHB inhibits the cleavage of caspase-1, which turns IL-1β into its active form in LPS-primed mouse macrophages and human monocytes. This effect is independent of fasting-regulated mechanisms known to act on the NLRP3 inflammasome. Moreover, the effect of BHB on caspase-1 activation and IL-1β secretion has been shown in mouse models of Muckle-Wells syndrome, a disorder associated with a pathogenic variant of the NLRP3 gene. Finally, KD and the associated elevation in BHB levels protected mice bearing the missense NLRP3 mutation that leads to familial cold autoinflammatory syndrome [72]. Furthermore, KD can reduce ROS production by promoting mitochondrial biogenesis [73]. ROS can also activate the NLRP3 inflammasome protein, triggering innate immune defenses through proinflammatory cytokines [74].
6. Other possible candidates
The IL-1–IL-1Ra axis is closely connected to the NLRP3 pathway [18]. Lin and Hsu [19] suggested potential therapeutics other than anakinra that might inhibit this pathway. Drugs targeting NLRP3 inflammasome assembly or activation include interferon-1, MCC950 (a highly specific small-molecule inhibitor for NLRP3), and proton pump inhibitors. A monoclonal anti-IL-1β antibody (canakinumab) and IL-1 blocker (rilonacept) can also be considered to block IL-1β activity [19].
Conclusion
Accumulating data support the proposal that the cytokine pathway mediated through IL-1β is involved in both human and animal models of epilepsy, including FIRES. Dysregulation of the IL-1β–IL-1Ra axis and inhibition affects epilepsy and epileptogenesis. These factors can be modified or targeted by therapeutics, including anakinra, which has evidence of potent anti-seizure effects in various experimental models of seizures, affords neuroprotection, and has potential anti-epileptogenic effects.
Notes
Joon Won Kang and Sookyong Koh are the editorial board members of the journal, but They was not involved in the peer reviewer selection, evaluation, or decision process of this article. No other potential conflicts of interest relevant to this article were reported.
Author contribution
Conceptualization: SK. Data curation: JWK. Formal analysis: JWK. Funding acquisition: JWK. Methodology: JWK. Project administration: JWK and SK. Visualization: JWK. Writing-original draft: JWK. Writing-review & editing: SK.
Acknowledgements
We thank Prof. Eun-Kyeong Jo for her comments on the figure. This work was supported by Chungnam National University.